
Pilzanbau/ Biologische Grundlagen
| Dieses Kapitel der biologischen Grundlagen der Pilze befindet sich derzeit im Prozess der Entstehung. Habe bitte Verständnis, dass der Hauptautor die Zügel in der Hand behalten will. Frag vor inhaltlichen Veränderungen daher bitte erst bei diesem an, um die Zusammenarbeit abzustimmen. Außerdem bittet dich der Hauptautor bei inhaltlichen Vorschlägen eine Referenz anzugeben, damit die Korrektheit geprüft werden kann. Korrekturen der Rechtschreibung sind natürlich immer gerne gesehen. Vielen Dank. |
Motivation
[Bearbeiten]Die Grundlagen der Biologie werden nur zu gern als unnötige Theorie empfunden, ist der Pilzanbau im kleinen Maßstab doch eher von praktischer Natur. Und so kommt es immer wieder zu Ratschlägen in unzähligen (auch gedruckten) Anleitungen, die jeden Erkenntnissen der Mykologie widersprechen. Wer die Theorie kennt, der befreit sich aus den Fesseln der Anleitung und muss sich bei der Aufzucht nicht mehr ausschließlich auf fremde Erfahrungen stützen. Früher oder später wird man beim Pilzanbau vor Problemen stehen, die man mit seiner Anleitung nicht bewältigen kann - wer aber weiß wie diese Lebewesen funktionieren, der kann nun seine eigenen Schlüsse ziehen, eine Lösung finden und daraus lernen. Ganz nebenbei wird man dabei vielleicht die Faszination der Biologie entdecken und bemerken wie nah Theorie und Praxis doch beieinander liegen können.
Alle Informationen in diesem Kapitel beruhen auf den Erfahrungen und Forschungen von erfahrenen und anerkannten Mykologen sowie Biologen. Die meisten der dabei konsultierten Quellen finden Sie im letzten Abschnitt mit einer kurzen Einschätzung zu Aufbau und Inhalt.
Einführung in das Reich der Pilze
[Bearbeiten]Die Organismengruppe der Pilze (Fungi) umfasst bisher über 75.000 bekannte Arten (man schätzt die gesamte Artenvielfalt auf etwa 1,5 Millionen) und steht taxonomisch gesehen mit den Pflanzen (Plantae) und Tieren (Animalia) auf einer Stufe. Pilze gehören damit also weder den Tieren noch den Pflanzen an. Von diesen drei Reichen (so nennt man diese Stufe der Unterteilung von Lebewesen) ist das der Pilze das unerforschteste, was sich für den Interessierten auch in der verfügbaren Literatur widerspiegelt. Von den vielen Pilzen sind für den Anbauer nur wenige von Interesse, hauptsächlich jene aus der Abteilung der Ständerpilze (Basidiomycota). Die meisten Speise- und viele Giftpilze die wir aus dem Alltag womöglich kennen, gehören zu dieser Gruppe. Aber selbst diese Abteilung umfasst immer noch rund 30.000 bekannte Arten, es fällt daher verständlicherweise schwer die Biologie dieser für eine kurze Einführung auf einen gemeinsamen Nenner zu bringen. Wir wollen es dennoch versuchen und begrenzen und dazu weiter auf die Unterabteilung der Agaricomycotina, diese Pilze bilden ein unterirdisches Pilzgeflecht aus, das sogenannte Myzel. Insbesondere wollen wir in dieser Unterabteilung die Klasse der Agaricomycetes betrachten, welche meist oberirdische Fruchtkörper ausbilden. Ob Champignon, Fliegenpilz, Psilocybe, Pfifferling oder Tintling - sie alle gehören dieser Klasse an.
Bedauerlicherweise sind nur die wenigsten von den nunmehr etwa 21.000 Arten erforscht. Wir werden uns daher im Folgenden häufig auf eine spezielle Art (Coprinopsis Cinerea) beschränken, wenngleich viele der Merkmale auch allgemeiner gültig sind.
Lebenszyklus
[Bearbeiten]
Häufig unterteilt man das Wachstum von Pflanzen und Pilzen in eine vegetative und reproduktive (oder generative) Phase. Während des vegetativen Wachstums bilden sich dabei die Organe aus, die dem Lebewesen der Ernährung dienen. Bei den hier betrachteten Pilzen handelt es sich dabei um das meist unterirdische Myzelgeflecht. In der reproduktiven Phase hingegen bilden Pilze häufig Fruchtkörper aus die mit den Sporen besetzt sind. Wir wollen diese Unterteilung auch in dieses Kapitel als grobe Einteilung übernehmen, wenn auch die beiden Phasen zeitlich nicht (oder nicht immer) voneinander zu trennen sind.
Die vegetative Phase
[Bearbeiten]Keimung
[Bearbeiten]Die Sporen der meisten Ständerpilze sind einkernig und haploid. Einkernig bedeutet dass die Sporen jeweils nur einen Zellkern besitzen und haploid, dass sich in jedem Zellkern nur ein einfacher Chromosomensatz befindet. Es existiert also in jedem Zellkern nur eine einfache Kopie eines jeden Chromosom. Das Myzel eines Stammes kann viele Fruchtkörper hervorbringen und jeder davon bildet in seiner Entwicklung unzählige Sporen. Unter geeigneten Umweltbedingungen keimt nun eine solche Spore und es entwächst eine Hyphe - der Pilzfaden. Die Hyphe besteht aus aneinandergereihten länglichen Zellen (genauer: zellähnlichen Gebilden), getrennt durch Querwände (Septen).
Primärmyzel
[Bearbeiten]In jeder dieser, durch die Septen getrennten, Zellen befindet sich (in der Regel) genau ein Zellkern. Die letzte (endständige) Zelle der Hyphe ist jene, die sich verdoppelt und damit den Pilzfaden wächsen lässt. Jeder der Kerne der Hyphe ist also ein genetisches Ebenbild des Kernes der vorhergehenden Zelle und damit letztlich eine Kopie des Kerns in der Spore. Man bezeichnet diesen Zustand, in dem alle Zellkerne in den einzelnen Zellen genetische Kopien sind (also vom ein und dem selben Genotyp sind), als homokaryotisch. Sollte sich in jeder Zelle noch dazu nur ein einzelner Zellkern befinden (was nicht immer der Fall sein muss), so bezeichnet man den Organimus oder einen Abschnitt des selben als homokaryotisches Monokaryon oder auch einfach als Monokaryon (in der Regel impliziert monokaryotisch auch homokaryotisch). Diese erste Form des Myzels wird Primärmyzel genannt. Kurz nach der Keimung befinden sich in den Zellen der jungen Hyphe oft noch viele gleiche Zellkerne, die erst ein wenig später durch Querwände (Septen) getrennt werden. Es ist also zu Beginn homokaryotisch, aber erst nach der Trennung monokaryotisch.
Sekundärmyzel
[Bearbeiten]Anastomose dikaryotischer Hyphen
Die Anzahl der Sporen die ein Pilz freigibt, ist so hoch, dass praktisch in allen natürlichen Fällen mehr als nur eine davon keimt. So dass diese monokaryotischen Pilzfäden schließlich aufeinander treffen können. Die beiden Hyphen bilden in diesem Fall eine Querverbindung untereinander aus (Anastomose), und sollten beide Hyphen vegetativ kompatibel sein, so wandert einer der beiden Zellkerne aus seiner in die andere Zelle über. Falls nun zusätzlich die beiden Kerne auch noch vom selben Kreuzungstyp sein sollten, so bilden sich fortan - während des weiteren Wachstums - Zellen mit jeweils zwei Kernen. Bei vielen Pilzarten werden auch die älteren Abschnitte der Hyphe nachträglich noch zweikernig. Wichtig ist dass beide Kerne noch nicht miteinander verschmelzen!
Was genau man unter der vegetativen Kompatibilität und dem Kreuzungstyp versteht, wird im Kapitel zur Genetik näher erläutert. Es sei aber hier noch kurz erwähnt dass, sollten die beiden Hyphen nicht kompatibel sein, die verbundenen Zellen absterben um das restliche Myzel zu schützen.
Die neue zweikernige Form des Myzels wird häufig als Sekundärmyzel bezeichnet. Da sich nun in jeder (fortan) wachsenden Zelle zwei Kerne befinden statt einem, ist es nicht mehr mono- sondern dikaryotisch. Außerdem befinden sich nun in den Zellen nicht mehr nur Zellkerne eines Genotyps sondern je einer von jeder der verschmolzenen Hyphen, dieser Zustand heißt daher auch nicht mehr homo- sondern heterokaryotisch. Erst ab dieser Phase ist das Myzel theoretisch in der Lage, Fruchtkörper zu bilden. Ob das Sekundärmyzel jedoch tatsächlich in der Lage ist diese auszubilden, hängt vom Kreuzungstyp und anderen genetischen Faktoren ab. So gibt es auch heterokaryotisches Myzel, das weiterhin nicht im Besitz der dazu benötigten Mechanismen ist (siehe Abschnitt Genetik).
Dikaryotisches Myzel unterscheidet sich unter dem Mikroskop (ab ca. 100-400 facher Vergrößerung) häufig - aber nicht immer - durch das Auftreten so genannter Schnallen (englisch clamp connections). Diese bilden sich nur entlang einer dikaryotischen Hyphe und erinnern in ihrer Form tatsächlich an Heftklammern. Die erzeugten Ausbuchtungen benötigt die Zelle während der Kernteilung, um einen der beiden Kerne aus der neuen Zelle temporär auszulagern und anschließend in die Ausgangs-Zelle zu transportieren (siehe Bild). Makroskopisch gesehen unterscheiden sich mono- und dikaryotisches Myzel vor allem in der Wachstumsrate, wobei dikaryotisches Myzel dem monokaryotischen überlegen ist.
-
 Hyphe mit Schnalle
Hyphe mit Schnalle -
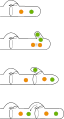 Entstehung der Schnalle
Entstehung der Schnalle


Für verschiedene Arten wurde gezeigt, dass der Pilz für das vegetative Wachstum (das Wachstum der Organe die der Ernährung des Organismus dienen - also des Myzels) absolute Dunkelheit bevorzugt - sowohl mono- aber vor allem dikaryotisches Myzel wird durch Beleuchtung in seinem Wachstum gehemmt.
Die reproduktive Phase (am Beispiel des C. Cinerea)
[Bearbeiten]
Sobald das Myzel die Oberfläche erreicht, fängt es unter geeigneten Umweltbedingungen an, Fruchtkörper zu bilden - auch diese bestehen weiterhin aus Hyphen die sich bis in die Lamellen (sofern es sich um einen Blätterpilz handelt) erstrecken. Innerhalb des Fruchtkörpers nehmen die Hyphen unterschiedliche Formen und Funktionen an, es handelt sich also um eine Art Spezialisierung (englisch hyphal differentiation). Wie genau diese abläuft und welche Funktion die einzelnen Arten einnehmen ist noch immer recht unerforscht und soll hier daher auch nicht Teil der Einführung sein. Selbst der grobe Ablauf wie sich ein Fruchtkörper entwickelt, unterscheidet sich je nach Art stark und selbst aus ein und dem selben Myzel können vollkommen unterschiedliche Formen von Fruchtkörpern entwachsen (z.B. durch Umwelteinflüsse).
Um dennoch eine Ahnung von den Entwicklungsphasen zu vermitteln werden wir uns im Folgenden auf eine spezielle Art beschränken. Der Zucht-Champignon, d.h. der zweisporige Egerling (Agaricus bisporus) ist den meisten Lesern in Form und vermutlich auch Geschmack am ehesten vertraut. Leider unterscheidet sich dieser in der Art seiner Sporen stark vom Großteil der Ständerpilze, weshalb er als allgemeines Beispiel nur bedingt brauchbar ist.
Weniger bekannt, dafür jedoch wissenschaftlich am stärksten erforscht ist der wollstielige Mist-Tintling (Coprinopsis Cinerea). Er entspricht mit seinem Stiel, dem Hut und den Lamellen dem typischen Bild eines Pilzes und ähnelt auch in der Sporenbildung vielen anderen Ständerpilzen, weshalb wir uns auf diese Spezies zur Veranschaulichung einigen wollen.

Man unterscheidet die Fruchtkörper und deren Entwicklung nach vielen Kriterien. Ein Kriterium ist etwa ob es ein Velum gibt das die Fruchtschicht (Hymenium), also den Ort an dem die Sporen gebildet werden, schützt und ob und ggf. wann in der Entwicklung dieses schützende Haut verschwindet. Der hier betrachtete Pilz C. Cinerea gehört dabei zu jenen Pilzen, deren Fruchtschicht (d.h. die Lamellen) vom Velum geschützt wird bis die Sporen reif sind. Danach löst sich das Velum und die Sporen können ungehindert ihren Weg vom von den Lamellen des Pilzes weg finden. Bei dieser Art der Entwicklung spricht man von Hemiangiocarp, sie unterscheidet sich von Pilzen bei denen die Fruchtschicht stets ungeschützt ist (Gymnocarp) oder auch jenen bei denen das Hymenium auch nach der Reifung weiterhin geschützt bleibt (Angiocarp).
Primärer Hyphen-Knoten (primary hyphal knot)
[Bearbeiten]Die Fruchtung beginnt mit einer extrem kleinen (ca. 0,1mm großen) Ballung von Hyphen. Diese besteht aus einer oder einigen wenigen Hyphen, die lokal vermehrt Verzweigungen bilden und so eine Art Knoten bilden. Hier beginnt bereits die erste Differenzierung der Hyphen: Eine oder mehrere Haupthyphen die die Richtung vorgeben und viele kurze Abzweiger die aus teilweise räumlich aufgeblähten "Zellen" bestehen. Die Hyphen sind in diesem Stadium noch nicht verschmolzen.
Sekundärer Hyphen-Knoten (secondary hyphal knot)
[Bearbeiten]Im weiteren Verlauf verzweigen die Hyphen immer mehr und schwellen weiterhin an. Die Hyphen fusionieren teilweise sogar miteinander (Anastomose). Während sich die Hyphen im Inneren des Knotens weiterhin stark ähneln, bildet sich um den jungen Fruchtkörper eine Art frühes Velum universale (eine Hülle um den gesamten Pilz). Im oberen Teil des Knotens beginnt nun die Differenzierung in Hut und Stiel. Der sekundäre Knoten ist in etwa 0,2mm im Durchmesser.
Primordium
[Bearbeiten]Entwicklungsstadien P1-P5 nach Kües
Fruchtkörper
[Bearbeiten]V.a. Bildung neuer Sporen, Verschmelzung von Kernen!
Genetik
[Bearbeiten]Chromosomenen, Chromatiden, Gene und Allele, Kreuzungstypen, vegetative Kompatibilität, Mitose, Meiose, zufällige Mutationen, "Geschlechter" der Sporen
Erscheinungsformen des Myzel
[Bearbeiten]Das Myzel kann in unterschiedlichen Formen auftreten, man unterscheidet für gewöhnlich zwischen rizomorphen (strangartigen) und watteähnlichen Myzel sowie einer Zwischenform. Vor allem bei der Petrischalenkultur ist die Sektorbildung besonders auffällig. Als Sektor wird dabei ein Teil des Myzels bezeichnet, der sich vom restlichen unterscheidet. Beispiel: Ein watteähnlicher Sektor in einem rizomorphen Myzelgeflecht.
Watteähnliches Myzel in der Petrischale führt dabei häufig zum sogenannten schwimmenden Myzel auf der Deckerde. Diese Erscheinungsform des Myzels ist besonders dicht und hemmt meist die Fruchtkörperbildung. Aus diesem Grund selektiert der Pilzbauer in der Regel die watteähnliche Erscheinungsform aus und züchtet lediglich die rizomorphe Form in seinen Kulturen weiter.
Die Entstehung und Funktion der unterschiedlichen Formen ist nicht abschließend geklärt. Es wurde aber gezeigt (z.B. G. Fritsche in "Flauschbildung beim Kulturchampignon" - Theoretical and Applied Genetics 40, 1970), dass auch der Nährboden neben der Genetik und dem Alter des Myzels erheblichen Einfluss auf die Flauschbildung hat.
Glossar
[Bearbeiten]Einige grundlegende biologische Begriffe häufen sich in den obigen Erläuterungen und bedürfen daher einer kurzen Definition. Wenngleich die Begriffe im Textfluss ebenfalls erläutert wurden, so vereinfacht sich das Verständnis doch durch eine gesammelte Auflistung zum nachschlagen:
- monokaryotisch
- Man spricht von einem Monokaryon, wenn jede Zelle genau einen Zellkern enthält.
- dikaryotisch
- Befinden sich genau zwei Zellkerne in jeder Zelle, so handelt es sich um ein Dikaryon. Die zwei Kerne sind in der Regel genetisch unterschiedlich, es ist aber keine Voraussetzung um dikaryotisch zu sein.
- homokaryotisch
- Wenn alle Zellkerne genetisch identisch sind (also vom selben Genotyp), so handelt es sich um ein Homokaryon.
- heterokaryotisch
- Es gibt zwei oder mehr genetisch unterschiedliche Zellkerne (unterschiedliche Genotypen).
- haploid (n)
- Ein Zellkern/eine Zelle der/die von jedem Chromosom genau eine Ausführung enthält (ein Chromosomensatz). Ein haploider Organismus besteht nur aus haploiden Zellen.
- diploid (2n)
- Siehe haploid, jedoch mit genau zwei Chromosomensätzen.
Weiterführende Literatur
[Bearbeiten]- Ursula Kües, Mónica Navarro-González, 2015, How do Agaricomycetes shape their fruiting bodies?
Ausgesprochen lehrreicher Artikel der aktuelle Erkenntnisse am Beispiel der drei wohl wichtigsten Modell- und Zucht-Pilze zusammenfasst. - Steven L. Stephenson, 2012, The Kingdom Fungi: The Biology of Mushrooms, Molds, and Lichens
- J. Stamets, 1984, The Mushroom Cultivator
Behandelt vor allem die Praxis der Kultivierung, beschränkt sich dabei leider teilweise auf Arten der Gattung Psilocybe. Sehr verständlich mit kürzeren Kapiteln zu den biologischen Hintergründen. Eine gute Einführung. - D. Moore, G. Robson, A. Trinci, 2011, 21st Century Guidebook to fungi
Ein sehr aktuelles und umfangreiches englischsprachiges Werk mit vielen Referenzen (an Ende eines jeden Kapitels) zu weiterführender Literatur. Behandelt vor allem die Biologie der Pilze. Sehr verständlich beschrieben und zum Selbststudium hervorragend geeignet. - J. Webster, 1983, Pilze: Eine Einführung
Eines der wenigen deutschsprachigen Bücher. Taxonomisch untergliedert. Erfordert teilweise grundlegende Kenntnisse der Biologie. - A. C. Chang, 1992, Genetics and Breeding of Edible Mushrooms
Ebenfalls sehr empfehlenswert da auch (relativ) aktuell. Verbindet die Biologie mit der Zucht in angenehmer Weise. - K. Esser, R. Kuenen, 1967, Genetik der Pilze
Sehr genau und ausführlich. Umfangreiche Vorkenntnisse notwendig und veraltet, für den Hobby-Mykologen daher oft nur bedingt brauchbar. - P. Miles, S. Chang, 1997, Mushroom Biology: Concise Basics and Current Developments
Gute und relativ aktuelle Einführung. Verständlich geschrieben.